
Types de mémoire : Concepts à la recherche
La mémoire est la fonction cognitive qui nous permet d’encoder, de stocker et de récupérer des informations. C'est une fonction essentielle à notre connaissance du monde et à notre fonctionnement quotidien. Dans cet article, nous passerons en revue les différents types de mémoire tout en discutant d’exemples de recherches et de tâches afin d’obtenir une compréhension pratique de la manière dont des expériences sont menées pour découvrir davantage sur les différents types de mémoire.
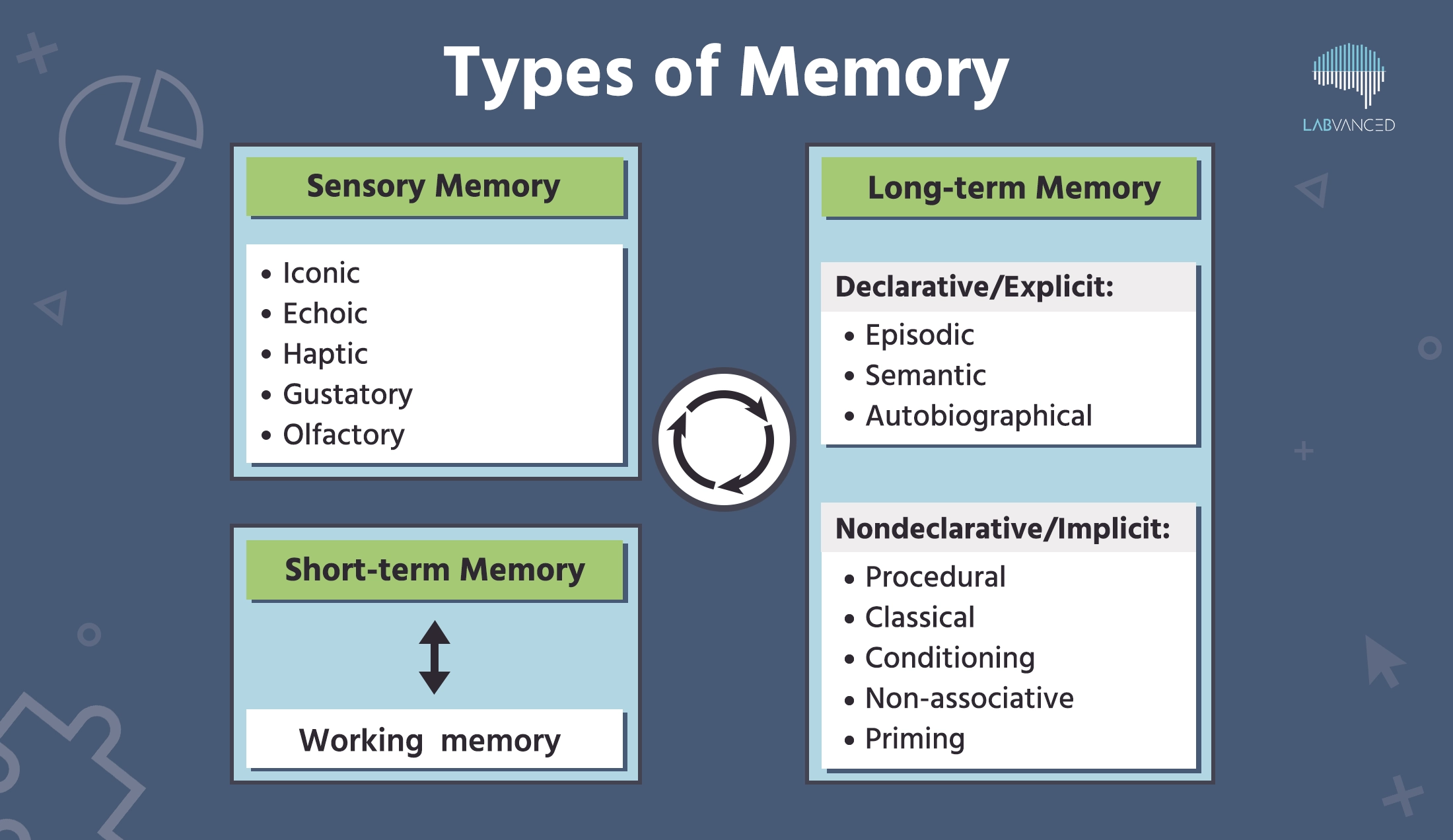
La mémoire est typiquement catégorisée en mémoire sensorielle, mémoire à court terme et mémoire à long terme, chacune ayant ses propres caractéristiques distinctes. Plongeons-nous !
Mémoire sensorielle
Tout le processus de se souvenir commence avec l'information entrant dans la mémoire sensorielle, c'est-à-dire ce qui est capté par les cinq sens. Lorsque nous rencontrons des informations sensorielles (comme le son, le toucher, l'odorat), un bref stockage de celles-ci se produit et cela s'appelle la mémoire sensorielle. Celles-ci sont généralement de courte durée. Si l'attention est portée à cette information sensorielle, elle est encodée dans la mémoire à court terme (Ciolek & Lee, 2020).
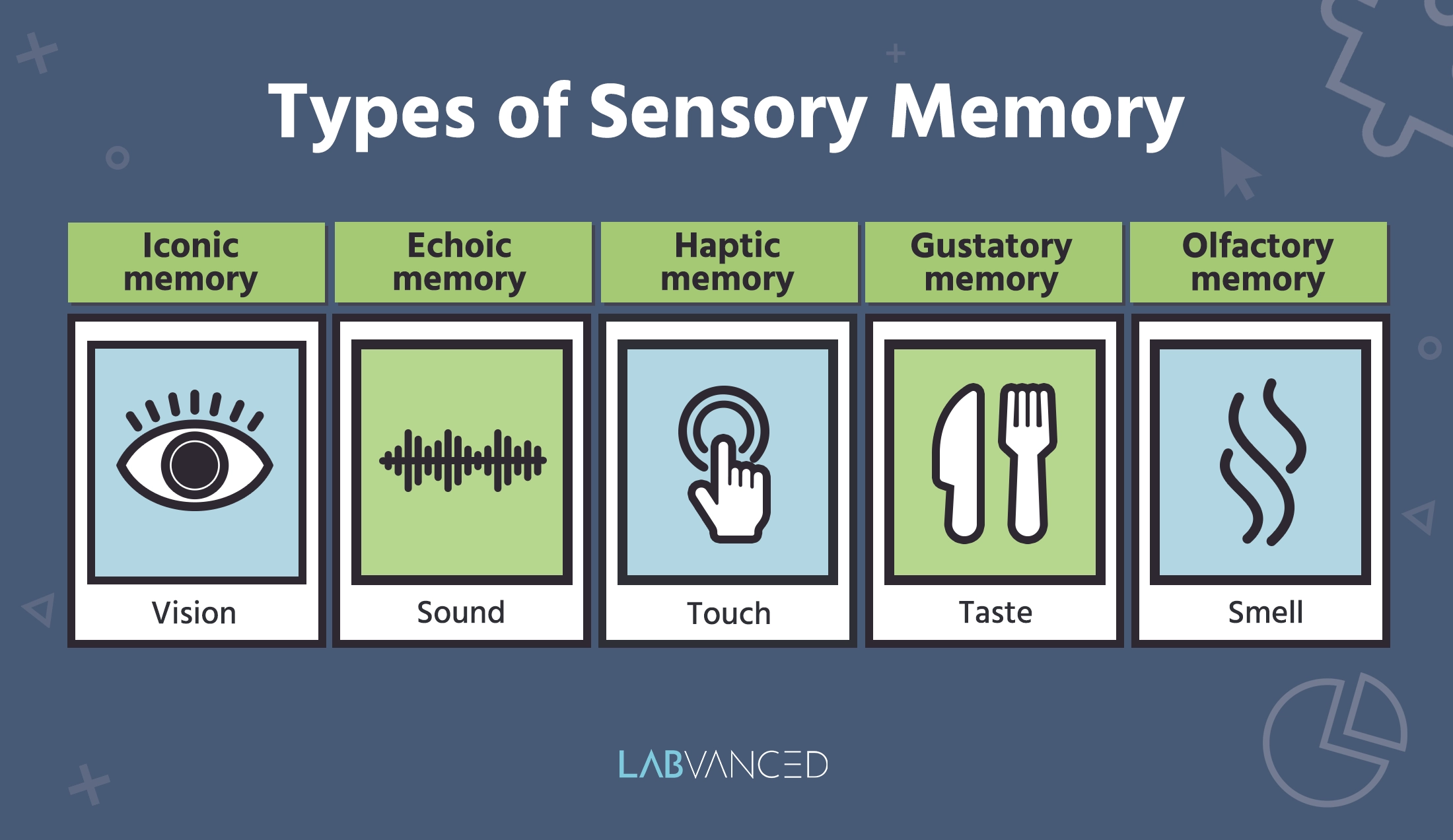
Finalement, des aspects de la mémoire sensorielle entreront dans la mémoire à long terme, vous permettant de chérir des souvenirs de choses comme le rythme de votre chanson blues préférée. Plongeons dans les types de mémoire sensorielle :
Mémoire iconique
La mémoire iconique est un sous-ensemble de la mémoire sensorielle qui est associé aux données visuelles, décrivant spécifiquement la rétention à court terme et la dégradation rapide des informations visuelles. Une personne ayant une mémoire iconique peut se souvenir d'une image d'une scène ou d'un objet pendant une très brève période de temps, généralement autour de 250 ms (Kanwar et al., 2023). Le mot "iconique" est dérivé du mot grec "eikōn" (εἰκών) signifiant “image”, soulignant ce composant visuel de ce type de mémoire (Klyukanov & Sinekopova, 2016).
L'expérience emblématique de George Sperling des années 1960, le paradigme du rapport partiel, est un excellent exemple de mémoire iconique. Dans cette expérience, une grille de lettres a été montrée aux participants pendant une courte période. Après que l'affichage a disparu, les participants ont été invités à rappeler soit l'ensemble de la grille, soit une ligne particulière de lettres. Sperling a découvert que lorsqu'on les incitait juste après l'affichage, les participants pouvaient rappeler avec précision l'ensemble de la grille, indiquant un stockage transitoire de l'information visuelle (Sperling, 1960).
Dans cet exemple ci-dessous dans Labvanced, les participants sont invités à saisir les chiffres qu'ils ont vu à l'écran dans une tâche de mémoire de chiffres :
Importez la tâche de mémoire de chiffres dans votre compte ou essayez-la simplement dans Labvanced.
Mémoire échoïque
La mémoire sensorielle à court terme connue sous le nom de « mémoire échoïque » est ce qui permet au système auditif de conserver des sons et d'autres stimuli pendant une courte période après que le stimulus initial s’est terminé. La mémoire échoïque persiste plus longtemps que la mémoire iconique et dure environ 1 à 2 secondes. La mémoire échoïque joue un rôle clé dans le traitement et la compréhension du langage parlé. Elle facilite le maintien d’un flux ininterrompu d’informations auditives, permettant aux personnes de percevoir et de comprendre le son, la musique ou d'autres types de stimuli auditifs (Kanwar et al., 2023 ; Cope et al., 2023).
Le phénomène connu sous le nom "d'effet de masquage inversé" est une illustration bien connue de la mémoire échoïque en action. Lorsqu'une personne entend une série de sons puis immédiatement après, un autre son (un masque) est présenté, cela peut obstruer leur perception et leur mémoire du premier son. Même si la mémoire échoïque est de courte durée, elle est essentielle à notre compréhension et à notre interprétation de notre environnement auditif (Edelman & Moyal, 2017).
Mémoire haptique
Le terme "mémoire haptique" décrit généralement un type de mémoire sensorielle associée aux haptics, ou au sens du toucher. Plus précisément, la mémoire haptique est la capacité de se souvenir des expériences liées au toucher. La mémoire haptique est un composant vital de notre expérience sensorielle globale, car elle nous aide à percevoir et à comprendre le monde physique qui nous entoure. De l'identification des textures à la manipulation d'objets, elle est impliquée dans de nombreuses tâches quotidiennes (Shihet al., 2009b).
📌 Point Publication : Le rôle du toucher affectif dans la promotion de l’attention des nourrissons vers des scènes visuelles complexes
Une étude de Carnevali, L., Della Longa, L., Dragovic, D., & Farroni, T. (2024) dans Labvanced a exploré le rôle du toucher affectif dans l'amélioration de l'attention des nourrissons vers des scènes visuelles complexes, mettant en évidence l'intégration des informations multisensorielles dans le développement précoce. L'étude basée sur Labvanced a utilisé :
- Conception de la Tâche : Une tâche expérimentale en trois phases pour évaluer l'impact du toucher affectif et non affectif sur l'attention des nourrissons à des stimuli visuels.
- Matériaux : Personnages animés (carré et triangle) associés à des stimuli auditifs (note musicale) et tactiles (toucher de main) pour créer des expériences multisensorielles engageantes.
- Collecte de Données : i) Les temps de regard des nourrissons ont été enregistrés lors des phases de familiarisation et de présentation visuelle pour mesurer l'attention et l'engagement ; ii) Des enregistrements vidéo des sessions ont permis le codage hors ligne des réponses des nourrissons.
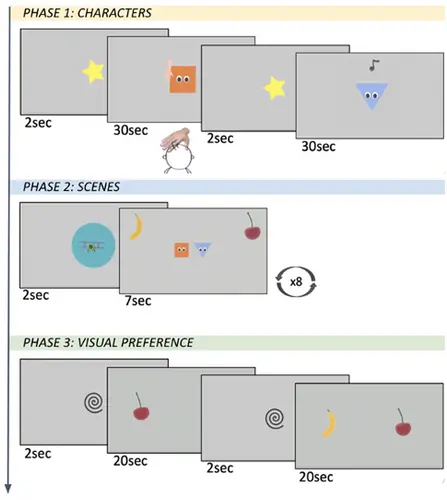
Résultats : Les chercheurs ont découvert que le toucher affectif (dans la phase 1 de l'expérience) menait à des temps de regard plus longs pendant la présentation des scènes (phase 2). Cela montre comment l'expérience ou la mémoire haptique peut jouer un rôle significatif dans la modulation de l'attention visuelle des nourrissons envers des scènes visuelles complexes en les prédisposant à explorer leur environnement et à faciliter une attention soutenue.
Mémoire gustative
La mémoire gustative se réfère à la mémoire du goût. La capacité de se souvenir d'un goût alimentaire spécifique est pertinente pour l'intake alimentaire, mais aussi pour l'apprentissage associatif. En recherche en psychologie, la mémoire gustative est généralement étudiée dans le contexte de la nourriture ou de l'aversion au goût et comment le goût ou les aliments déjà consommés guident les comportements futurs liés à l'alimentation (Lim et al., 2022).
Mémoire olfactive
La mémoire olfactive se réfère à la mémoire de l'odeur. Dans le monde naturel et dans le règne animal, l'odorat joue un rôle important dans la conduite du comportement, aidant à la survie, et même en aidant à la navigation spatiale (Yang et al., 2021). Une tendance récente en recherche en psychologie est d'étudier l'entraînement olfactif, c'est-à-dire l'entraînement à l'odorat avec une exposition répétée aux olfactifs dans le but de favoriser la neuroplasticité. Cela ouvre la voie à la compréhension de la façon dont la mémoire olfactive s'entrelace avec d'autres processus cognitifs, mais aussi en ce qui concerne des conditions pathologiques (Vance et al. ,2024).
📌 Point Publication : Compréhension du langage lié aux odeurs dans l'anosmie acquise
Dans cette étude menée dans Labvanced, les chercheurs ont voulu évaluer si l'anosmie acquise (la perte du sens de l'odorat qui survient plus tard dans la vie) impacte la compréhension du langage lié aux odeurs. Les chercheurs ont administré une série de tâches, afin d'obtenir un tableau complet de la mémoire liée aux odeurs chez les anosmiques et les contrôles. Fait intéressant, l'étude a conclu qu'il n'y avait aucune preuve que l'anosmie acquise nuise à la compréhension des mots liés aux odeurs ou aux goûts, mais que les associations émotionnelles avec les mots liés aux odeurs et aux goûts étaient altérées chez les anosmiques, avec des évaluations plus positives. Dans l'ensemble, ces résultats suggèrent que le traitement du langage peut dans certains cas être indépendant de la capacité à avoir une expérience sensorielle olfactive (Speed, L. J., Iravani, B., Lundström, J. N., & Majid, A., 2022).
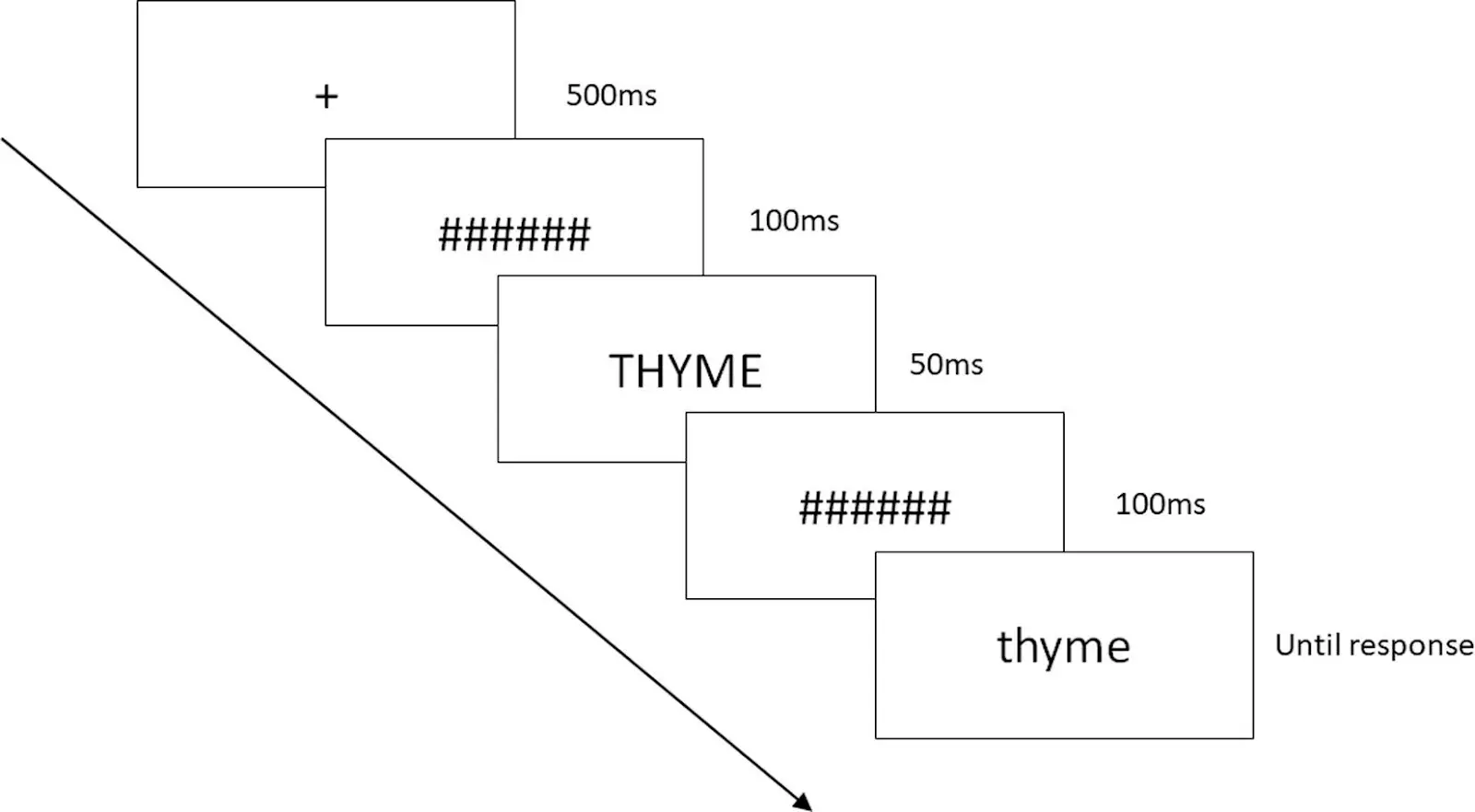
Exemple de la procédure de la tâche de décision lexicale réalisée dans Labvanced ; Speed, L. J., Iravani, B., Lundström, J. N., & Majid, A. (2022).
Mémoire à court terme
La mémoire à court terme (MCT) fait référence au traitement de petites quantités d'informations pendant une courte période. Lorsque nous rappelons une information immédiatement après qu'elle nous a été présentée, c'est la MCT en action ! En l'absence de répétition, l'information subit une dégradation rapide et dure donc moins de ~30 secondes. La MCT inclut la mémoire de travail, qui aide au processus de stockage de la mémoire à court terme (Camina & Güell, 2017 ; Kramer & Stephens, 2014).
Mémoire de travail
La mémoire de travail est responsable de la manipulation et du stockage temporaire d'informations. C'est la capacité de conserver des informations en tête pendant une courte période afin de pouvoir les utiliser pour un traitement supplémentaire. Pour des tâches cognitives impliquant le traitement simultané d’informations, comme suivre des instructions, résoudre des problèmes difficiles et comprendre un langage parlé ou écrit, la mémoire de travail est cruciale (Maricle & Bauman Johnson, 2016).
L'un des tests les plus courants utilisés pour mesurer la mémoire à court terme et la mémoire de travail visuospatiales est le test de frappe de blocs de Corsi (CBT) (Schaefer et al., 2022).
L'exemple ci-dessous montre le test de frappe de blocs de Corsi administré dans Labvanced :
Importez la tâche de frappe de blocs de Corsi dans votre compte pour votre prochaine expérience ou essayez-la simplement dans Labvanced.
Mémoire à long terme
Lorsque l'information stockée dans la mémoire à court terme se transfère vers le stockage à long terme, cela s'appelle la mémoire à long terme (MLT) et peut durer des jours, voire des années. La MLT implique les processus d'encodage, de stockage et de récupération d'informations (Johnson, R. (2014). Elle a été divisée en deux grandes catégories : la mémoire explicite et implicite.
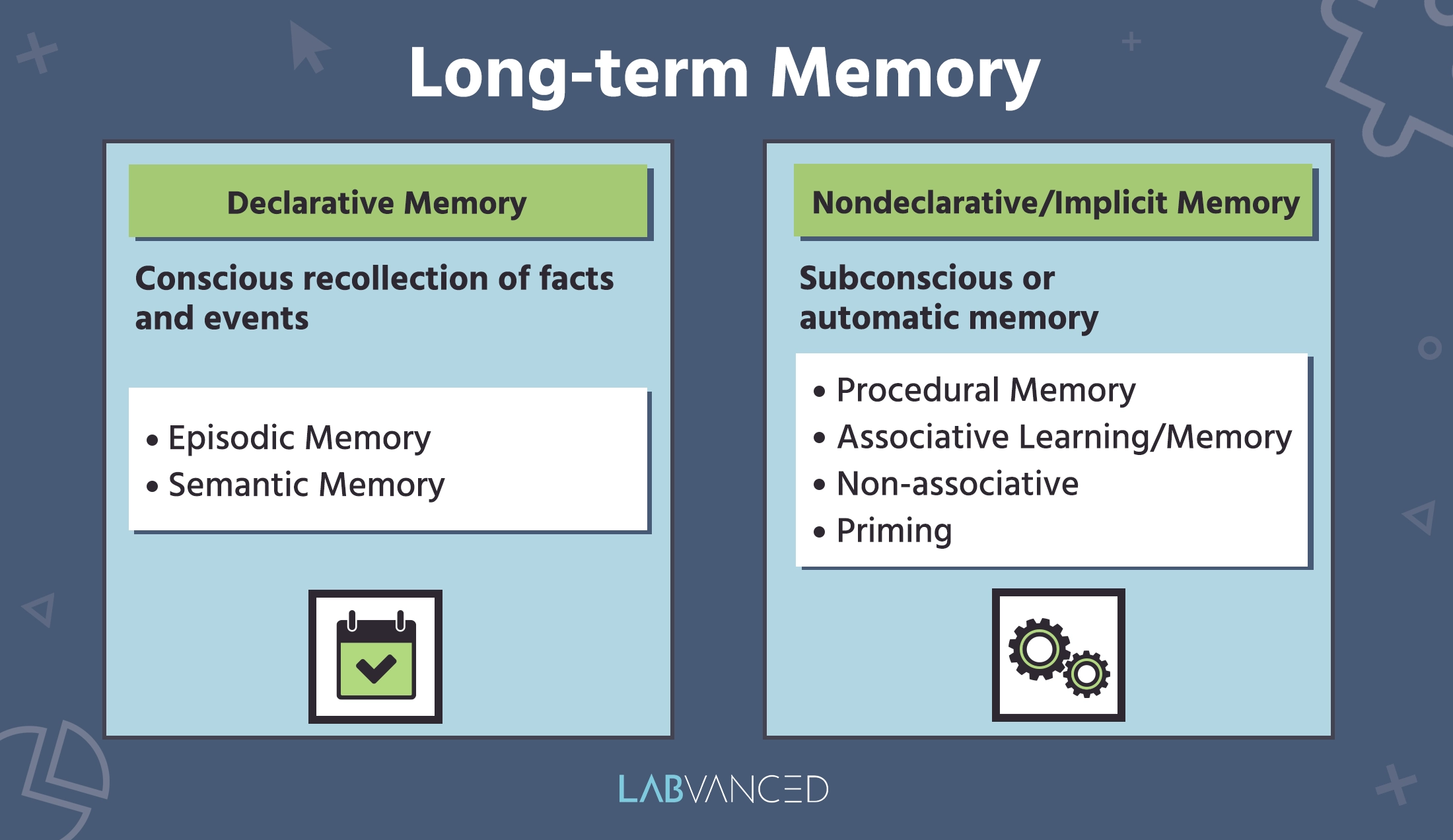
Mémoire déclarative / explicite :
La mémoire explicite, ou mémoire déclarative, est la catégorie de mémoire qui inclut le rappel délibéré d’événements et de faits. Elle se divise en deux types :
Mémoire épisodique
Un sous-ensemble de la mémoire déclarative qui traite des expériences personnelles et a la capacité de se souvenir d'expériences ou d'événements particuliers du passé d'une personne. C'est le souvenir des rencontres individuelles liées à un lieu et à un moment spécifiques.
La mémoire épisodique est souvent appelée ‘mémoire autobiographique.’ Par exemple, se souvenir de détails sur des événements, y compris ce qui s'est passé, où cela s'est passé, quand cela s'est passé et les sentiments ou émotions qui y étaient liés. La mémoire épisodique contient des détails sensoriels (détails ou informations perçues par les cinq sens) qui nous permettent de revivre une expérience (Morè et al., 2020).

Mémoire sémantique / mémoire autobiographique
C'est un autre sous-ensemble de la mémoire déclarative à long terme qui inclut des faits et des connaissances générales. Elle couvre une vaste gamme d’idées, telles que la langue, la réalité et les significations des mots. La mémoire sémantique est essentiellement l'ensemble des connaissances acquises que l'on a rassemblées sur le monde (Binder, J. R., & Desai, R. H., 2011). Cela inclut des choses comme les noms des couleurs, la compréhension du langage, ou les capitales des grandes villes.
La mémoire sémantique est plus abstraite et générale que la mémoire épisodique et n'est pas liée à des événements personnels spécifiques. Comparé à la mémoire épisodique, qui est la mémoire d'expériences spécifiques, la capacité de la mémoire sémantique semble se développer plus tôt dans l'enfance. Des études montrent que la mémoire sémantique tend à se stabiliser avec le temps et même augmenter avec l'âge (Martin & Simmons, 2008 ; Richmond & Burnett, 2022).
Mémoire non déclarative / implicite
La mémoire implicite, un autre nom pour la mémoire non déclarative, est une catégorie de mémoire à long terme qui n’est pas rappelée consciemment. Ce type de mémoire inclut des réponses conditionnées, des compétences, des habitudes et des comportements. La plupart du temps, la mémoire non déclarative s'exprime et s'acquiert inconsciemment. Elle inclut également ce qui suit :
Mémoire procédurale
La mémoire procédurale est un sous-ensemble de la mémoire non déclarative ou implicite qui inclut la formation et le maintien de compétences et d'habitudes perceptivo-motrices, comme faire du vélo ou pratiquer un sport ! Elle permet aux gens d’agir et de prendre des décisions sans réaliser qu'ils le font ou avoir besoin d'évoquer des détails spécifiques de la mémoire. La mémoire procédurale joue un rôle critique dans notre capacité à apprendre et à développer des compétences motrices par la pratique et la répétition. En termes de neurobiologie, la mémoire procédurale implique des connexions plus fortes entre les voies synaptiques associées au comportement, résultant en des réponses réflexives et inconscientes (Fogel & Smith, 2011 ; Mayford et al., 2012).
Qu'est-ce que Labvanced ?
Labvanced est une plateforme puissante conçue spécifiquement pour réaliser des expériences comportementales et cognitives ainsi que des recherches psychologiques utilisant des fonctionnalités avancées telles que le suivi oculaire validé par des pairs et le support d'études multi-utilisateurs via des applications web et de bureau/mobile natives.
Dans une étude conduite dans Labvanced par Gavard, E. & Ziegler, J.C. (2024), une série de tâches a été mise en œuvre pour déterminer si l'apprentissage statistique (un mécanisme cognitif qui permet d'apprendre des motifs de manière implicite) pouvait indiquer des prédictions linguistiques pendant la lecture. Une tâche pertinente ici était la tâche de temps de réaction en série (SRT) qui est une tâche classique d'apprentissage moteur séquentiel. Les chercheurs ont administré cela afin d'obtenir une idée de l'apprentissage (statistique) implicite des participants, qui est considéré comme lié au système de mémoire procédurale.
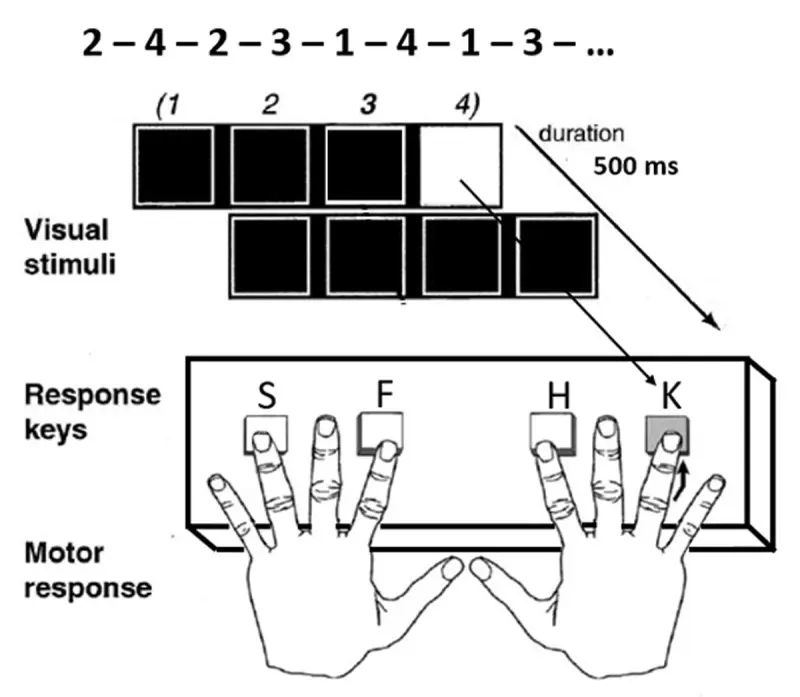
Conception expérimentale et timing de la tâche SRT (adaptée de Schendan et al., 2003) Gavard, E. & Ziegler, J. C. (2024) dans Labvanced.
Mémoire associative ou conditionnement classique
La mémoire associative est la capacité de reconnaître et de se souvenir des relations entre des objets apparemment non liés. Par exemple, développer une peur des chiens après avoir été mordu. Les individus possédant ce type de mémoire peuvent récupérer des connaissances davantage à travers des connexions qu'à partir d'indices ou de détails spécifiques. Par exemple, vous pourriez reconnaître quelqu'un que vous avez vu dans la rue grâce au parfum d'un magasin à proximité (Suzuki, 2008).
📌 Mise en avant de publication : Apprentissage et mémoire associés et symptômes de PTSD analogues
Friesen, E., et al (2022) ont cherché à déterminer si la détresse et la rumination liées à l'épidémie de COVID-19 pouvaient influencer le développement de symptômes analogues au PTSD lorsque les participants étaient exposés à un clip vidéo aversif non lié au COVID-19. Les chercheurs voulaient également examiner si cette relation était médiée par la force de l'apprentissage associatif (c'est-à-dire le conditionnement de la peur). Les chercheurs ont administré leur expérience et tâche d'apprentissage associatif en utilisant Labvanced. L'image ci-dessous montre la configuration expérimentale et la procédure de l'étude. Les résultats suggèrent qu'une détresse accrue pendant la pandémie de COVID-19 a pu renforcer l'apprentissage associatif, conduisant à une augmentation des souvenirs intrusifs et de la rumination en réponse à un événement traumatique analogue.
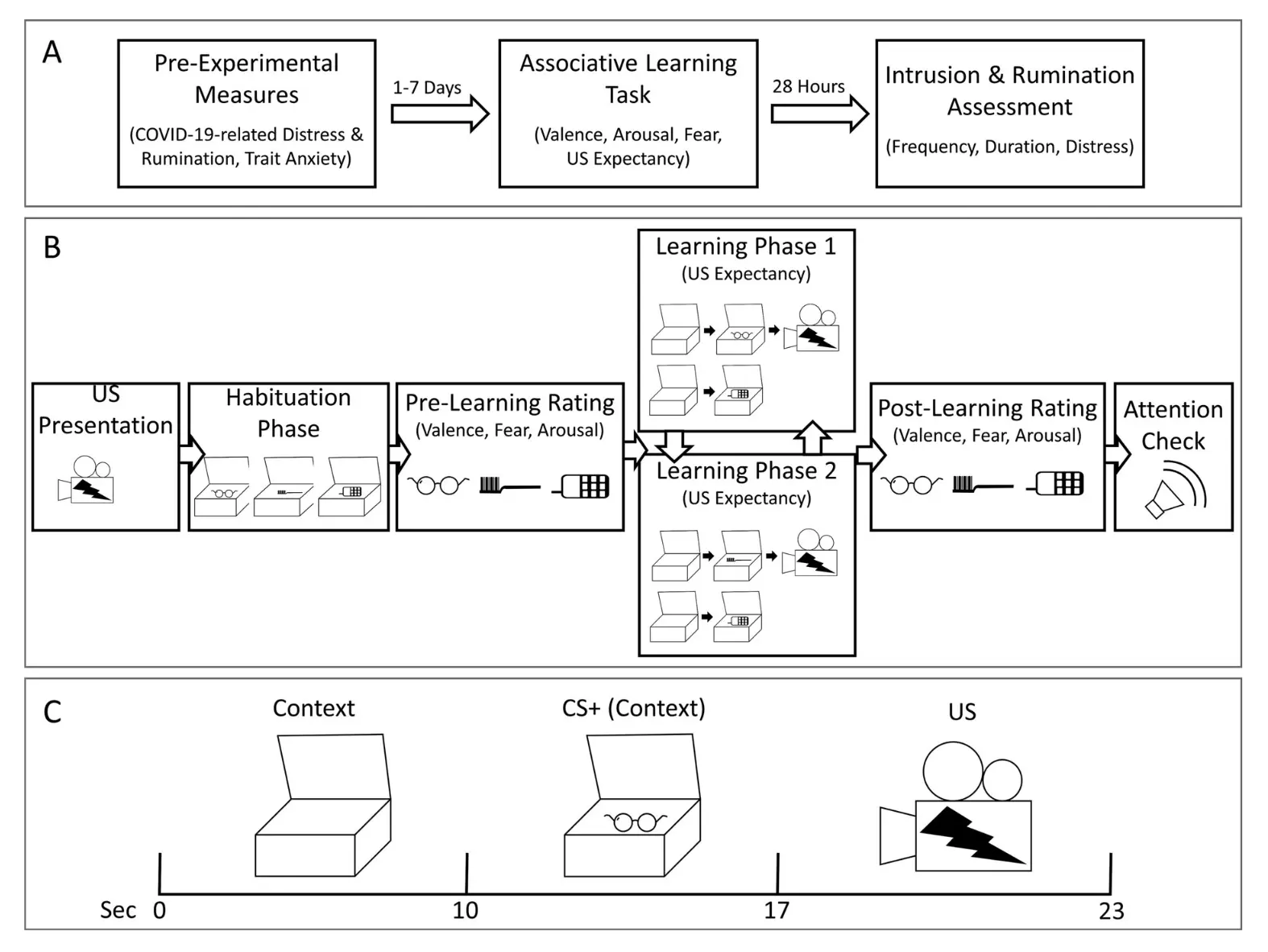
Illustration de la procédure d'étude réalisée dans Labvanced.
Remarque : (A) Procédure générale de l'étude. (B) Procédure de la tâche d'apprentissage associatif différentiel. (C) Présentation des stimuli lors d'un essai CS+ renforcé pendant la tâche d'apprentissage associatif différentiel. CS+ = stimulus conditionné ; US = stimulus non conditionné ; Friesen, E., et al (2022).
Mémoire non associative
La mémoire non associative, également connue sous le nom d'apprentissage non associatif, est un apprentissage qui ne nécessite pas d'association ou de couplage de stimuli. Ce style d'instruction est considéré comme l'approche la plus fondamentale et la plus simple car il repose sur un changement de comportement en réponse à un seul stimulus. Il met plus l'accent sur l'acquisition et la conservation de composants distincts sans nécessiter de connexions ou de liaisons entre eux (Ioannou & Anastassiou-Hadjicharalambous, 2018). Les sujets importants dans ce domaine incluent l'habituation et la sensibilisation. Un exemple d'apprentissage non associatif par habituation serait qu'après avoir vécu un certain temps dans une rue animée, vous finissez par « vous y habituer ».
Mémoire de priming
C'est un effet de mémoire implicite dans lequel l'exposition à un stimulus influence les réponses à un autre, appelé "priming" en psychologie. Cela implique l'activation de schémas particuliers dans la mémoire à long terme pour faciliter le rappel d'informations pertinentes. Par exemple, lorsqu'un individu est exposé à la couleur jaune et qu'on lui demande ensuite de nommer rapidement un fruit, il est plus probable qu'il dise le mot "banane" plutôt que "pomme" ou "raisin". Cette technique de priming est appliquée en psychologie pour enseigner aux gens comment réagir à certains stimuli ou modifier leur comportement. Sans que la personne ne le comprenne, le priming influence la fonction cognitive de manière consciente ou inconsciente. Comprendre comment une exposition préalable à des stimuli peut affecter le comportement et les processus cognitifs dans le présent est facilité par le priming (Bermeitinger, 2015).
Dans l'image ci-dessous, un ensemble échantillon de stimuli d'une étude menée par Baumann, L., & Valuch, C. (2022) dans Labvanced, montre les nombreux stimuli utilisés pour étudier l'effet du traitement sémantique et du priming dans une tâche de catégorisation. Les participants étaient invités à catégoriser l'image prime (ignorant la cible) comme étant soit une scène intérieure, soit extérieure. La prime était présentée très brièvement (50 ms), suivie de la cible (400 ms maximum). Les chercheurs ont découvert que le temps de réaction était significativement plus rapide lorsque la prime et la cible appartenaient à la même catégorie congruente (c'est-à-dire que les deux étaient des scènes extérieures).

Ensemble échantillon de stimuli utilisés dans Labvanced pour étudier le priming de la catégorisation des scènes.
(A-gauche) Exemples de stimuli cibles de chaque contexte de la catégorie extérieure (A-droite) catégorie intérieure (B) Image du haut -gauche : exemple de stimuli prime pour la catégorie extérieure ; images du bas -gauche : stimuli cibles pour la catégorie extérieure pour les terrains de jeux ; Image du haut -droite : exemple de stimuli prime pour la catégorie intérieure ; images du bas -droite : stimuli cibles pour la catégorie extérieure pour la cuisine ;
Baumann, L., & Valuch, C. (2022).
Conclusion
La mémoire est un système complexe et multifacette. Comprendre ses formes et ses différents types nous aide à apprendre comment nous retenons, traitons et rappelons les informations, façonnant finalement notre apprentissage, notre perception, notre interaction et notre navigation dans la vie quotidienne dans le monde !
Références
Baumann, L., & Valuch, C. (2022). Priming of natural scene categorization during continuous flash suppression. Consciousness and Cognition, 104, 103387.
Bermeitinger, C. (2015). Priming. Advances in Psychology, Mental Health, and Behavioral Studies, 16–60.
Binder, J. R., & Desai, R. H. (2011). The neurobiology of semantic memory. Trends in cognitive sciences, 15(11), 527-536.
Camina, E., & Güell, F. (2017). The neuroanatomical, neurophysiological, and psychological basis of memory: Current models and their origins. Frontiers in Pharmacology, 8.
Chaaya, N., Battle, A. R., & Johnson, L. R. (2018). An update on contextual fear memory mechanisms: Transition between amygdala and hippocampus. Neuroscience & Biobehavioral Reviews, 92, 43–54.
Ciolek, C. H., & Lee, S. Y. (2020). Cognitive issues in the older adult. Guccione’s Geriatric Physical Therapy, 425–452.
Cope, T. E., Sohoglu, E., Peterson, K. A., Jones, P. S., Rua, C., Passamonti, L., Sedley, W., Post, B., Coebergh, J., Butler, C. R., Garrard, P., Abdel-Aziz, K., Husain, M., Griffiths, T. D., Patterson, K., Davis, M. H., & Rowe, J. B. (2023). Temporal lobe perceptual predictions for speech are instantiated in motor cortex and reconciled by inferior frontal cortex. Cell Reports, 42(5), 112422.
Edelman, S., & Moyal, R. (2017). Fundamental computational constraints on the time course of perception and action. Progress in Brain Research, 121–141.
Fogel, S. M., & Smith, C. T. (2011). The function of the sleep spindle: A physiological index of intelligence and a mechanism for sleep-dependent memory consolidation. Neuroscience & Biobehavioral Reviews, 35(5), 1154–1165.
Friesen, E., Michael, T., Schäfer, S. K., & Sopp, M. R. (2022). COVID-19-related distress is associated with analogue PTSD symptoms after exposure to an analogue stressor. European Journal of Psychotraumatology, 13(2), 2127185.
Gavard, E., & Ziegler, J. C. (2024). Semantic and syntactic predictions in reading aloud: Are good predictors good statistical learners?. Journal of Cognition, 7(1), 40.
Ioannou, A., & Anastassiou-Hadjicharalambous, X. (2018). Non-associative learning. Encyclopedia of Evolutionary Psychological Science, 1–13.
Johnson, R. (2014). The neural basis of deception and credibility assessment. Credibility Assessment, 217–300.
Kanwar, S., Bafna, G., & Gogania, P. (2023). Assessment of short-term memory by the word and object test in young adults. International Journal of Health Sciences and Research, 13(6), 90–92.
Klyukanov, I. E., & Sinekopova, G. (2016). Beyond the Binary: Toward the Paraconsistencies of Russian Communication Modes. International Journal of Communication, 10, 17.
Kramer, J. H., & Stephens, M. L. (2014). Memory, overview. Encyclopedia of the Neurological Sciences, 1045–1047.
Maricle, D. E., & Bauman Johnson, W. L. (2016). Instructional implications from the Woodcock–Johnson IV tests of cognitive abilities. WJ IV Clinical Use and Interpretation, 123–150.
Martin, A., & Simmons, W. K. (2008). Structural basis of semantic memory. Learning and Memory: A Comprehensive Reference, 113–130.
Mayford, M., Siegelbaum, S. A., & Kandel, E. R. (2012). Synapses and memory storage. Cold Spring Harbor Perspectives in Biology, 4(6).
Morè, L., Lauterborn, J. C., Papaleo, F., & Brambilla, R. (2020). Enhancing cognition through pharmacological and environmental interventions: Examples from preclinical models of neurodevelopmental disorders. Neuroscience & Biobehavioral Reviews, 110, 28–45.
Lim, S. X. L., Höchenberger, R., Ruda, I., Fink, G. R., Viswanathan, S., & Ohla, K. (2022). The capacity and organization of gustatory working memory. Scientific reports, 12(1), 8056.
Richmond, L. L., & Burnett, L. K. (2022). Characterizing older adults’ real-world memory function using ecologically valid approaches. Psychology of Learning and Motivation, 193–232.
Schaefer, S. Y., Hooyman, A., Haikalis, N. K., Essikpe, R., Lohse, K. R., Duff, K., & Wang, P. (2022). Efficacy of Corsi Block Tapping Task training for improving visuospatial skills: a non-randomized two-group study. Experimental Brain Research, 240(11), 3023-3032.
Schendan, H. E., Searl, M. M., Melrose, R. J., & Stern, C. E. (2003). An FMRI study of the role of the medial temporal lobe in implicit and explicit sequence learning. Neuron, 37(6), 1013-1025.
Shih, R., Dubrowski, A., & Carnahan, H. (2009a). Evidence for haptic memory. World Haptics 2009 - Third Joint EuroHaptics Conference and Symposium on Haptic Interfaces for Virtual Environment and Teleoperator Systems, 145–149.
Shih, R., Dubrowski, A., & Carnahan, H. (2009b). Evidence for haptic memory. World Haptics 2009 - Third Joint EuroHaptics Conference and Symposium on Haptic Interfaces for Virtual Environment and Teleoperator Systems, 145–149.
Speed, L. J., Iravani, B., Lundström, J. N., & Majid, A. (2022). Losing the sense of smell does not disrupt processing of odor words. Brain and language, 235, 105200.
Sperling, G. (1960). The information available in brief visual presentations. Psychological Monographs: General and Applied, 74(11), 1–29.
Suzuki, W. A. (2008). Chapter 19: Associative learning signals in the brain. Progress in Brain Research, 305–320.
Vance, D. E., Del Bene, V. A., Kamath, V., Frank, J. S., Billings, R., Cho, D. Y., ... & Fazeli, P. L. (2024). Does olfactory training improve brain function and cognition? A systematic review. Neuropsychology review, 34(1), 155-191.
Yang, A. I., Dikecligil, G. N., Jiang, H., Das, S. R., Stein, J. M., Schuele, S. U., ... & Gottfried, J. A. (2021). The what and when of olfactory working memory in humans. Current Biology, 31(20), 4499-4511.