
记忆类型:从概念到研究
记忆是我们编码、存储和检索信息的认知功能。它是我们对世界知识和日常功能的重要组成部分。在本文中,我们将讨论不同类型的记忆,并讨论研究和任务的示例,以便深入了解如何进行实验来发现不同类型的记忆。
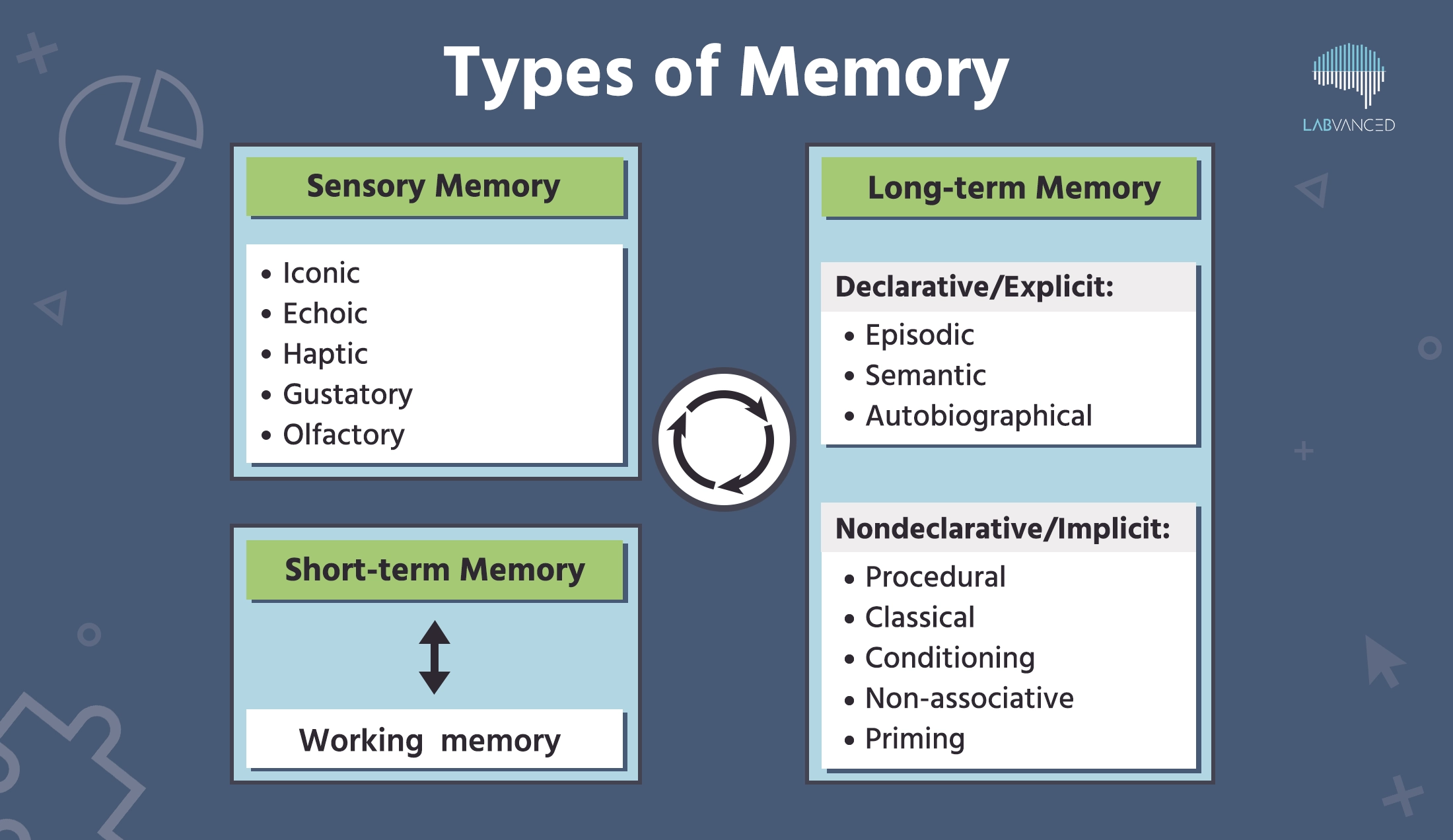
记忆通常被分为感觉记忆、短期记忆和长期记忆,每种记忆都有其独特的特征。让我们深入探讨吧!
感觉记忆
记忆的整个过程始于信息进入感觉记忆,即输入五感的信息。当我们遇到感觉信息(例如声音、触感、气味)时,会发生短暂的存储,这称为感觉记忆。这些通常是短暂的。如果对这些感觉信息给予注意,它就会被编码到短期记忆中(Ciolek & Lee, 2020)。
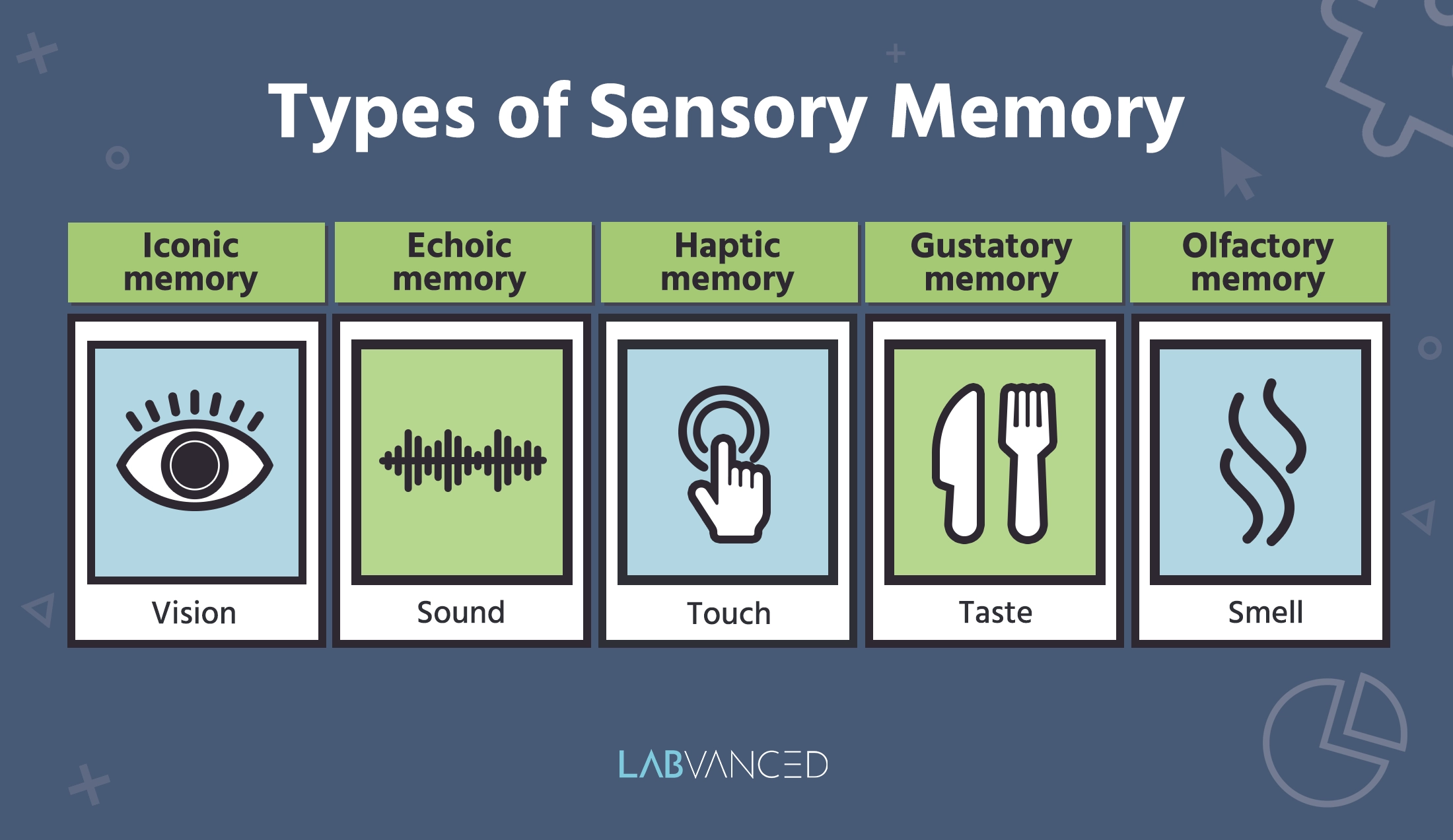
最终,感觉记忆的某些方面将进入长期记忆,使您能够珍藏像您最喜欢的布鲁斯歌曲的旋律这样的美好回忆。让我们深入了解不同的 感觉记忆类型:
图像记忆
图像记忆是感觉记忆的一个子集,与视觉数据相关,特别描述视觉信息的短期保持和快速衰退。具有图像记忆的人可以在非常短的时间内(通常约 250 毫秒)保留场景或物体的图像(Kanwar et al., 2023)。 "图像"一词源自希腊词 "eikōn" (εἰκών),意为“图像”,突显了这种记忆的视觉组成部分(Klyukanov & Sinekopova, 2016)。
乔治·斯佩林(George Sperling)在 1960 年代的开创性实验——部分报告范式,是图像记忆的一个很好的例子。在这个实验中,一组字母在一个短时间内呈现给参与者。在显示消失后,参与者被要求回忆整个字母表或特定的字母行。斯佩林发现,当在显示后立即提示时,参与者能够准确回忆整个字母表,这表明了视觉信息的瞬态存储(Sperling, 1960)。
下面的 Labvanced 示例中,参与者被要求在数字跨度任务中输入他们刚看到的数字:
将 数字跨度任务 导入到您的帐户中,或者在 Labvanced 中直接尝试。
回声记忆
被称为“回声记忆”的短期感觉记忆,使听觉系统能够在初始刺激结束后短时间内保留声音和其他刺激。回声记忆的持续时间比图像记忆长,通常持续约 1 到 2 秒。回声记忆在处理和理解语言方面扮演了重要角色。它有助于保持听觉信息的连续流,使人们能够感知和理解声音、音乐或其他类型的听觉刺激(Kanwar et al., 2023; Cope et al., 2023)。
“向后掩蔽效应”这一现象是回声记忆作用的一个著名例子。当一个人听到一系列声音,随后立刻呈现另一个声音(掩蔽物),可能会妨碍他们对第一个声音的感知和记忆。尽管回声记忆是短暂的,但它对我们理解和解释听觉环境至关重要(Edelman & Moyal, 2017)。
触觉记忆
“触觉记忆”通常描述与触觉相关的感觉记忆类型。具体来说,触觉记忆是记住触觉相关体验的能力。触觉记忆是我们整体感觉体验的重要组成部分,因为它帮助我们感知和理解我们周围的物理世界。从识别纹理到处理物体,它参与许多日常任务(Shihet al., 2009b)。
📌 出版物聚焦:情感触摸在促进婴儿对复杂视觉场景的注意力中的作用
一项由 Carnevali, L., Della Longa, L., Dragovic, D., & Farroni, T. (2024) 进行的 Labvanced 研究探讨了情感触摸在增强婴儿对复杂视觉场景的注意力中的作用,强调了在早期发展中多感觉信息整合的重要性。 该基于 Labvanced 的研究使用了:
- 任务设计: 三阶段实验任务,评估情感和非情感触摸对婴儿对视觉刺激注意力的影响。
- 材料: 动画角色(正方形和三角形)与听觉(音乐音符)和触觉(手触)刺激配对,以创建引人入胜的多感觉体验。
- 数据收集: i) 在熟悉和视觉展示阶段记录婴儿的注视时间,以测量注意力和参与度; ii) 对会议的视频录制便于对婴儿反应的离线编码。
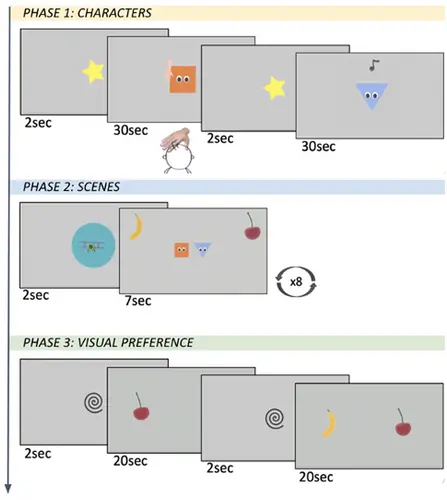
研究结果: 研究人员发现,在实验的第一阶段,情感触摸导致场景展示期间的注视时间更长(第二阶段)。这显示了触觉体验或记忆在调节婴儿对复杂视觉场景的视觉注意力方面可能发挥重要作用,让他们倾向于探索环境并促进持续注意力。
味觉记忆
味觉记忆是指对味道的记忆。记住特定食物味道的能力与食物摄入相关,同时也与 联想学习。 在心理学研究中,味觉记忆通常在食物或味道厌恶的背景下进行研究,以及味道或以前消费食物如何指导未来的饮食相关行为(Lim et al., 2022)。
嗅觉记忆
嗅觉记忆是指对气味或嗅觉的记忆。在自然界和动物王国中,嗅觉在指导行为、帮助生存甚至协助空间导航方面发挥着重要作用(Yang et al., 2021)。最近心理学研究的一个趋势是研究嗅觉训练,即通过多次接触气味进行的嗅觉训练,目的是实现神经可塑性。这为理解嗅觉记忆如何与其他认知过程交织在一起开辟了道路,同时也为病理状况提供了帮助(Vance et al., 2024)。
📌 出版物聚焦:获得性嗅觉丧失对嗅觉相关语言理解的影响
在 Labvanced 进行的这项研究中,研究人员希望评估获得性嗅觉丧失(指在生命后期失去嗅觉)是否会影响对嗅觉相关语言的理解。研究人员实施了一系列任务,以全面了解嗅觉丧失者和对照组的嗅觉相关记忆。有趣的是,该研究得出的结论是,没有证据表明获得性嗅觉丧失会损害对嗅觉或味道词的理解,但嗅觉丧失者与嗅觉和味道词的情感关联有所改变,评估较为积极。总体而言,这些研究结果表明,在某些情况下,语言处理可能与嗅觉感觉体验的能力无关(Speed, L. J., Iravani, B., Lundström, J. N., & Majid, A., 2022)。
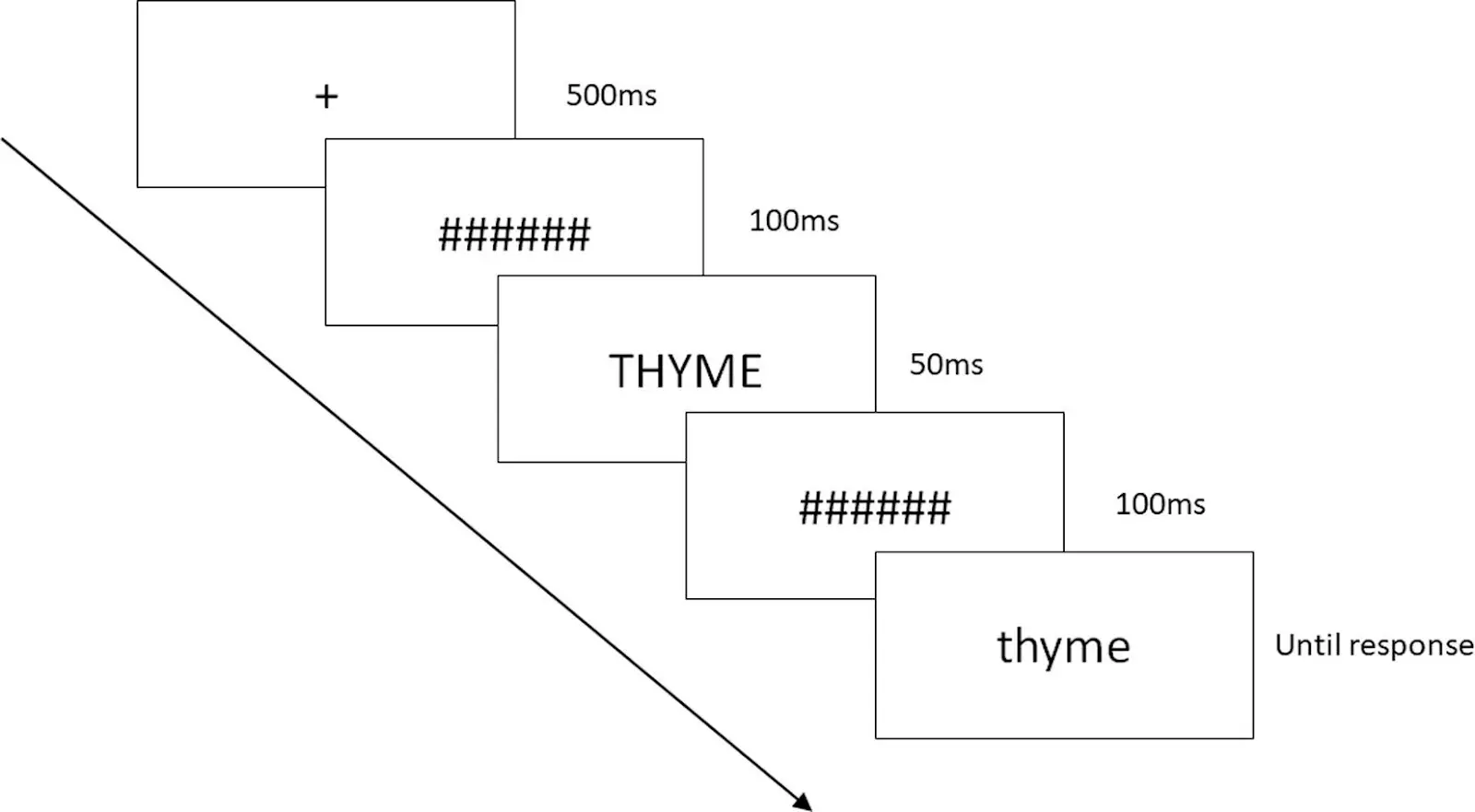
在 Labvanced 中进行的词汇决策任务程序示例; Speed, L. J., Iravani, B., Lundström, J. N., & Majid, A., (2022).
短期记忆
短期记忆(STM)是指对少量信息进行短时间处理的能力。当我们在信息呈现后立即回忆起某些信息时,这就是短期记忆的表现!如果没有复述,这些信息会迅速消逝,通常持续不到约 30 秒。短期记忆还包括工作记忆,有助于执行短期记忆存储的过程(Camina & Güell, 2017; Kramer & Stephens, 2014)。
工作记忆
工作记忆负责操纵和暂时存储信息。它是将信息保留在脑海中一段短时间,以便您可以用它进行进一步处理的能力。对于涉及信息同时处理的认知任务,如跟随指示、解决困难问题以及理解口头或书面语言,工作记忆至关重要(Maricle & Bauman Johnson, 2016)。
用于测量视觉空间短期和工作记忆的最常用测试之一是 Corsi 块敲击测试 (CBT)(Schaefer et al., 2022)。
下面的示例展示了在 Labvanced 中施行的 Corsi 块敲击测试:
将 Corsi 块敲击任务 导入到您的帐户以便进行下次实验,或在 Labvanced 中直接尝试。
长期记忆
当储存在短期记忆中的信息转移到长期存储时,就称为长期记忆(LTM),持续时间可以长达数天甚至数年。长期记忆包括信息的编码、存储和检索过程(Johnson, R. (2014))。它已被分为两个广泛的类别——显性和隐性记忆。
陈述性 / 显性记忆:
显性记忆或陈述性记忆是一个包括有意识回忆事件和事实的记忆类别。它分为两种类型:
事件记忆
显性记忆的一个子集,涉及个人经历,能够回忆起一个人过去的特定经历或事件。它是与特定地点和时间相关的个别经历的回忆。
事件记忆通常被称为「自传性记忆」。例如,回忆某些事件的细节,包括发生了什么、发生在哪里、何时发生以及与之相关的任何感受或情绪。事件记忆包含通过五感感知的感觉细节(细节或信息),让我们重温经历(Morè et al., 2020)。

语义记忆 / 自传性记忆
这是显性长期记忆的另一个子集,包括事实和一般知识。它涵盖了大量的概念,例如语言、现实和单词含义。语义记忆本质上是个人积累的关于世界的所有知识(Binder, J. R., & Desai, R. H., 2011)。这包括诸如颜色名称、语言理解或主要城市的首都等内容。
与事件记忆相比,语义记忆更加抽象和一般化,且与特定个人经历无关。与针对特定经历的事件记忆相比,语义记忆的能力在儿童时期就显示出发展的早期趋势。研究表明,语义记忆有稳定的趋势,并且随着年龄的增长而增加(Martin & Simmons, 2008; Richmond & Burnett, 2022)。
非陈述性 / 隐性记忆
隐性记忆,非陈述性记忆的另一种说法,是一种不被有意识回忆的长期记忆类。此类记忆包含条件反应、技能、习惯和行为。隐性记忆通常是以无意识的方式表现和获取的。它进一步包括以下几种:
程序性记忆
程序性记忆是非陈述性或隐性记忆的一个子集,包括感知-运动技能和习惯的形成与维持,如骑自行车或运动!它负责使人们能够在不意识到或不需要回忆特定细节的情况下采取行动和做出决策。程序性记忆在人们通过练习和重复学习和发展运动能力的能力中发挥着关键作用。在神经生物学方面,程序性记忆涉及与行为相关的突触通路之间更强的连接,导致反射和无意识的反应(Fogel & Smith, 2011; Mayford et al., 2012)。
什么是 Labvanced?
Labvanced 是一个强大的平台,专门用于进行行为和认知实验以及心理学研究,具有高级功能,如同行评审的眼动追踪和通过网络及本地桌面/移动应用程序的多用户研究支持。
在 Labvanced 中进行的一项研究,由 Gavard, E. & Ziegler, J.C. (2024) 进行,实施了一系列任务,以确定统计学习(使得模式可以隐含学习的认知机制)是否能够在阅读时指示语言预测。这里的一个相关任务是序列反应时间任务(SRT),这是一个经典的顺序运动学习任务。研究人员实施这一任务以了解参与者的隐含(统计)学习,这被认为与程序性记忆系统有关。
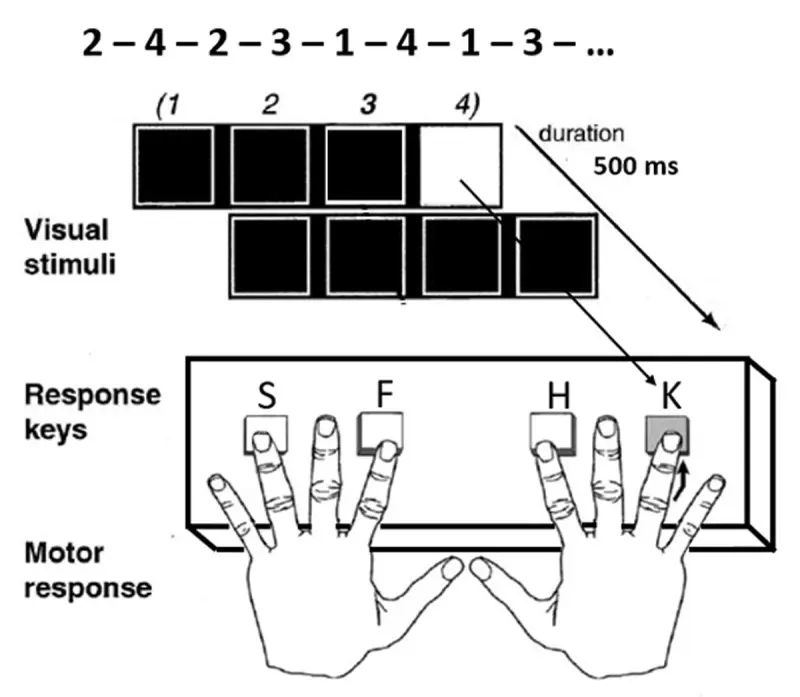
实验设计与 SRT 任务的时序(改编自 Schendan et al., 2003) Gavard, E. & Ziegler, J. C. (2024) 在 Labvanced。
联想记忆或经典条件反射
联想记忆是识别和记住看似不相关物体之间关系的能力。例如,在被狗咬后产生对狗的恐惧。拥有这种记忆类型的人能够通过连接而不是特定提示或细节来检索知识。例如,您可能会通过附近购物中心的香水的气味认出您在街上见过的某个人(Suzuki, 2008)。
📌 出版物亮点:联想学习和记忆与类似 PTSD 症状
Friesen, E. 等 (2022) 旨在确定 COVID-19 大流行相关的痛苦和反刍是否会影响在参与者暴露于非 COVID-19 相关的厌恶影片片段后类似 PTSD 症状的发展。研究人员还希望调查这种关系是否由联想学习的强度(即恐惧条件反射)所介导。研究人员使用 Labvanced 进行了实验及联想学习任务。下面的图像显示了实验设置和研究过程。研究结果表明,COVID-19 大流行期间的情绪高涨可能加强了联想学习,从而导致对类似创伤事件的侵入性记忆和反刍增加。
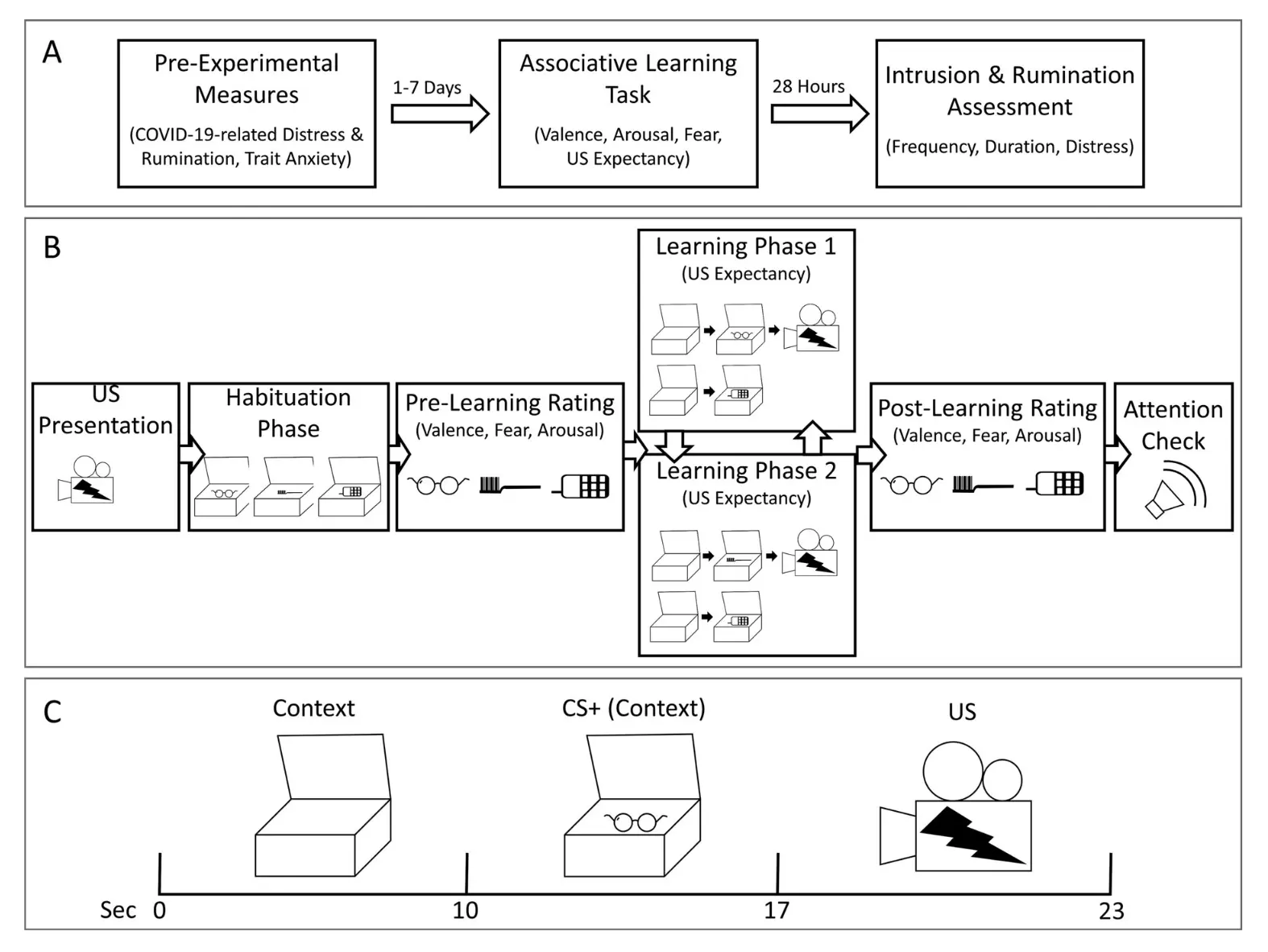
在 Labvanced 进行的研究程序示意图。
注意:(A) 一般研究程序。(B) 差异联想学习任务的程序。(C) 在差异联想学习任务中的强化 CS+ 试验中的刺激呈现。CS+ = 条件刺激;US = 无条件刺激; Friesen, E., et al (2022)。
非联想记忆
非联想记忆,也被称为非联想学习,是不需要刺激关联或配对的学习。这种教学风格被认为是最基本和最简单的方法,因为它依赖于对单一刺激的反应而引起的行为变化。它更加重视获取和保持单独成分,而不需要它们之间的连接。该领域的重要主题包括习惯化和敏感化。例如,对习惯化的非联想学习一例就是,住在繁忙街道一段时间后,您最终对其“习惯”。
诱导记忆
这是一种隐性记忆效应,其中对一个刺激的接触影响对另一个刺激的反应,心理学上称之为“诱导”。它涉及激活长期记忆中特定的图式,以促进相关信息的检索。例如,当一个人接触到黄色并随后被问到快速命名一种水果时,他们更可能说出“香蕉”而不是“苹果”或“葡萄”。在心理学中,诱导技术被应用于教导人们如何对特定刺激做出反应或改变他们的行为。在没有意识的情况下,诱导影响认知功能,无论是有意识的还是无意识的。诱导的帮助有助于理解先前对刺激的接触可能如何影响现在的行为和认知过程(Bermeitinger, 2015)。
在下面的图像中,来自 Labvanced 进行的 Baumann, L. 和 Valuch, C. (2022) 研究中的样本刺激集,展示了用于研究语义处理和分类任务中诱导效应的众多刺激。参与者被要求将诱导图像(忽略目标)分类为室内或室外场景。诱导短暂呈现(50 毫秒),随之而来的是目标(最大 400 毫秒)。研究人员发现,当诱导和目标属于同一一致类别(即都是户外场景)时,反应时间显著更快。

用于研究场景分类诱导的刺激样本集。
(A-左) 户外类别的每个上下文的目标刺激示例 (A-右) 室内类别 (B) 上图 -左:户外类别的诱导刺激示例;下图 -左:户外类别的游乐场目标刺激; 上图 -右:室内类别的诱导刺激示例;下图 -右:户外类别的厨房目标刺激;
Baumann, L., & Valuch, C. (2022).
结论
记忆是一个复杂且多面的系统。理解它的形式和不同类型帮助我们了解如何保持、处理和检索信息,最终塑造我们对世界的学习、感知、互动和导航方式!
参考文献
Baumann, L., & Valuch, C. (2022). 自然场景分类的诱导。知觉与认知, 104, 103387.
Bermeitinger, C. (2015). 诱导。心理学、心理健康和行为研究的进展, 16–60.
Binder, J. R., & Desai, R. H. (2011). 语义记忆的神经生物学。认知科学趋势, 15(11), 527-536.
Camina, E., & Güell, F. (2017). 记忆的神经解剖学、神经生理学和心理学基础:当前模型及其起源。药理学前沿, 8.
Chaaya, N., Battle, A. R., & Johnson, L. R. (2018). 关于情境恐惧记忆机制的更新:杏仁核与海马体之间的转变。神经科学与生物行为评论, 92, 43–54.
Ciolek, C. H., & Lee, S. Y. (2020). 老年人的认知问题。Guccione 的老年人物理疗法, 425–452.
Cope, T. E., Sohoglu, E., Peterson, K. A., Jones, P. S., Rua, C., Passamonti, L., Sedley, W., Post, B., Coebergh, J., Butler, C. R., Garrard, P., Abdel-Aziz, K., Husain, M., Griffiths, T. D., Patterson, K., Davis, M. H., & Rowe, J. B. (2023). 颞叶感知预测语言的过程在运动皮层中实现,并由下额皮层协调。细胞报告, 42(5), 112422.
Edelman, S., & Moyal, R. (2017). 感知与行动时间流程的基本计算约束。脑研究进展, 121–141.
Fogel, S. M., & Smith, C. T. (2011). 睡眠纺锤的功能:智力的生理指标以及睡眠依赖的记忆巩固机制。神经科学与生物行为评论, 35(5), 1154–1165.
Friesen, E., Michael, T., Schäfer, S. K., & Sopp, M. R. (2022). COVID-19 相关的痛苦与暴露于类创伤影片后出现的类似 PTSD 症状之间的关联。欧洲心理创伤学期刊, 13(2), 2127185.
Gavard, E., & Ziegler, J. C. (2024). 朗读中的语义和句法预测:好的预测者是否是好的统计学习者?认知杂志, 7(1), 40.
Ioannou, A., & Anastassiou-Hadjicharalambous, X. (2018). 非联想学习。进化心理科学百科全书, 1–13.
Johnson, R. (2014). 欺骗与可信度评估的神经基础。可信度评估, 217–300.
Kanwar, S., Bafna, G., & Gogania, P. (2023). 通过单词和物体测试评估年轻人的短期记忆。国际健康科学与研究期刊, 13(6), 90–92.
Klyukanov, I. E., & Sinekopova, G. (2016). 超越二元性:俄罗斯沟通模式的多重一致性。国际传播期刊, 10, 17.
Kramer, J. H., & Stephens, M. L. (2014). 记忆概况。神经科学百科全书, 1045–1047.
Maricle, D. E., & Bauman Johnson, W. L. (2016). 来自 Woodcock–Johnson IV 认知能力测试的教育意义。WJ IV 临床使用与解释, 123–150.
Martin, A., & Simmons, W. K. (2008). 语义记忆的结构基础。《学习与记忆:综合参考》,113–130.
Mayford, M., Siegelbaum, S. A., & Kandel, E. R. (2012). 突触与记忆存储。《冷泉港生物学前沿》,4(6).
Morè, L., Lauterborn, J. C., Papaleo, F., & Brambilla, R. (2020). 通过药物和环境干预来增强认知:来自神经发育障碍的临床前模型的示例。神经科学与生物行为评论, 110, 28–45.
Lim, S. X. L., Höchenberger, R., Ruda, I., Fink, G. R., Viswanathan, S., & Ohla, K. (2022). 味觉工作记忆的容量和组织。科学报告, 12(1), 8056.
Richmond, L. L., & Burnett, L. K. (2022). 使用生态有效的方法表征老年人的真实记忆功能。学习与动机心理学, 193–232.
Schaefer, S. Y., Hooyman, A., Haikalis, N. K., Essikpe, R., Lohse, K. R., Duff, K., & Wang, P. (2022). Corsi 块敲击任务训练在改善视觉空间技能方面的有效性:一项非随机的双组研究。实验脑研究, 240(11), 3023-3032.
Schendan, H. E., Searl, M. M., Melrose, R. J., & Stern, C. E. (2003). 一项关于内侧颞叶在隐性和显性序列学习中作用的 FMRI 研究。神经元, 37(6), 1013-1025.
Shih, R., Dubrowski, A., & Carnahan, H. (2009a). 触觉记忆的证据。世界触觉 2009 - 第三届共同 EuroHaptics 会议和虚拟环境与遥控操纵系统的触觉接口研讨会, 145–149.
Shih, R., Dubrowski, A., & Carnahan, H. (2009b). 触觉记忆的证据。世界触觉 2009 - 第三届共同 EuroHaptics 会议和虚拟环境与遥控操纵系统的触觉接口研讨会, 145–149.
Speed, L. J., Iravani, B., Lundström, J. N., & Majid, A. (2022). 失去嗅觉并不会干扰对气味词的处理。大脑与语言, 235, 105200.
Sperling, G. (1960). 短暂视觉呈现中可用信息的研究。心理学专论:一般与应用, 74(11), 1–29.
Suzuki, W. A. (2008). 第19章:大脑中的联想学习信号。脑研究进展, 305–320.
Vance, D. E., Del Bene, V. A., Kamath, V., Frank, J. S., Billings, R., Cho, D. Y., ... & Fazeli, P. L. (2024). 嗅觉训练能改善大脑功能和认知吗?一项系统评估。神经心理学评论, 34(1), 155-191.
Yang, A. I., Dikecligil, G. N., Jiang, H., Das, S. R., Stein, J. M., Schuele, S. U., ... & Gottfried, J. A. (2021). 人类嗅觉工作记忆的内容和时间。当前生物学, 31(20), 4499-4511.